
Mostly Natural
GRIZZLIES
Of the Northern Rocky Mountains
For everything on the politics and policy of grizzly bear conservation
go to Grizzly Times

For everything on the ecology of Ursus arctos, go to All Grizzly


Temporal Patterns of Cutworm moths & Bears
Back in 1955 John Chapman published a paper in the journal Ecology describing a peculiar feeding activity by bears in the Mission Mountains of western Montana. Grizzlies and black bears were both rummaging through alpine talus fields—eating something. As it turned out, they were slurping up army cutworm moths that had concentrated there during the summer to feed on nectar of high-elevation flowers. Twenty plus years later John Craighead and his colleagues described grizzly bears gobbling up cutworm moths from under overturned talus rock in alpine reaches of the Scapegoat Mountains roughly 50 miles east of the Missions. Barring a few published descriptions by early mountaineers, these were the first and—for a while—the only written descriptions of bears feeding on cutworm moths. Since then, bear consumption of cutworm moths has been described at multiple sites in Glacier National Park and Absaroka Mountains of the Yellowstone ecosystem , with evidence of an explosive increase in this activity in the Absarokas since the mid-1980s. Its not clear whether this trend towards ever-more documented exploitation of moths by bears is real or simply an artifact of search effort and search image.

Annual Trends: The Absaroka Mountains of Greater Yellowstone


D
The figures and map at right show trends in observed use of moth sites by grizzly bears in the Absaroka Mountains in relation to summer climate (C) and the expanding distribution of Yellowstone's grizzly bears (D). Figure B is the centerpiece. It shows the number of sites where bears have been annually observed feeding on moths (dark turquoise dots), with trend accentuated by a dark-burgundy line representing a 3-year running average. The lighter dots and red line below show total number of bears observed on all sites any given year. The takeaways from this graph are, first, that observed use of moth sites increased from nil during the mid-1980s to an approximate asymptote around 2002, and, second, that around 2012 numbers of observed bears skyrocketed, but with all of this increase concentrated on the same number of sites. I have little doubt that this latter marked increase in bear foraging on moths reflects compensation for widespread losses of whitebark pine during 2002-2009 (see Whitebark Pine Trends). The top figure (A) shows the proportion of sites that hosted the first observed sightings of females with cubs-of-the-year during any given year--increasing to a peak during the mid-2000s, decreasing, and now increasing again. I will return to this trend and its significance later.
The longer-term trends shown in (B) beg for an explanation. The most glib is that researchers and other observers of relevance only got on to an already widespread phenomenon during the mid-1980s, and then got more motivated and efficient to/at finding moth sites thereafter. Not likely. I was involved in grizzly bear investigations throughout the 1980s, and involved with the first observation of bears consuming moths in the Absarokas. We had radio-monitored bears in many areas that later supported moth-feeding sites throughout the late 1970s and early 1980s, with no indication that any of these bears camped on moth sites during late summer and early fall. In fact, the first bear that we knew to use a moth site had shown no sign of the behavior during any of the previous years she had been collared. More could be said regarding the evidence of radio-marked bears, but the bottom line is all available evidence suggests that the burgeoning of moth-feeding among Yellowstone grizzly bears during the late-1980s and early 1990s was real.
If true, then this emergence of moth-feeding by grizzly bears as a real phenomenon also begs an explanation. Old guys who outfitted big-game hunters in the region during the 1940s and 1950s claim to remember bears on talus slopes that we currently know to be moth sites. There is also the evidence of bleached mule and horse bones near several moth sites, testimony to hunters out to kill bears during decades past. Prior to the 1970s, hunters killed both grizzly and black bears in Wyoming by luring them into baits, including old broken-down equids that were led into the mountains and shot. In other words, the bleached bones are testimony to this sordid practice on or near moth sites, which is mute testimony, in turn, to the presence of bears feeding on moths.
If grizzly bears were feeding on moths in the Absaroka Mountains during the 1940s, 50s, and earlier, then why would they disappear from these sites during the 60s and 70s? The answer can only be speculate, but potentially comprised of three parts. Moth populations could have been at a nadir because of weather and the advent of more effective insecticides (see Introduction). Moths are affected by both, and harmed by unusually wet weather, which was more common during the 60s than during the previous or following decade. Grizzlies that consumed moths might have been killed by hunters at a higher rate than bears engaged in other activities simply because moth sites are so predictable and exposed. And that harvest was probably unsustainable. Data presented by John Craighead and his co-authors in a 1994 monograph on Yellowstone grizzlies suggests that unsustainable killing was occurring outside Yellowstone National Park. In addition, part of the increase in bear activity on moth sites during the late 1980s and ealry 1990s could simply be from grizzlies colonizing areas containing moth aggregations, which is what figure D, above, would suggest. Moth sites are shown as dusky green, and areas colonized during the 1980s and 90s by the outward arrows; total grizzly bear distribution as of 2000 is shown in green. The rapid colonization of new areas by grizzlies during these decades could have been caused both by the attraction of moth sites as well as by effects of wildfires during 1988 that burned over 1 million acres, shown in red. A fire-related effect would be consistent with the particularly rapid increase in moth site use after 1988, evident in the trends of figure B.
Finally, use of moth sites by grizzly bears in the Absarokas has obviously varied from one year to the next. Some of this variation might be attributable to observer effort, and some to variation in moth populations. But the drop in use during 1993 to essentially nothing is a stark anomaly, and warrants some sort of explanation. My speculation at this point is that the drop in use during this year, as well as some of the inter-annual variation that followed, was caused by summer temperatures in the alpine, shown in figure C, above. 1993 was typified by the coldest average July-August temperatures we've seen during the last 40 years, which may have delayed snow melt, alpine phenology, and, with that, the aggregation of moths in alpine environments. This interpretation is consistent with a negative correlation during other years between levels of bear feeding on moths and average summer temperatures (the red line in C).
Seasonal Trends

Moth phenology--as well as bear activity--exhibit pronounced and consistent seasonal trends. Beginning with moths, figures A and B at left show data collected by various researchers showing seasonal migrations of moths from the plains to the mountains and back again (see Introduction).
The top figure distills some pioneering work by Ken Pruess published in the 1960s. The red lines denote numbers of moths observed at a lowland site moving west towards the mountains, peaking in late June, and then back east to the plains, gradually increasing to a maximum during early September. The gray and green squiggly lines represent cumulative captures of moths during early summer headed west during different years, also by Pruess, but at a high-elevation site in the foothills of the Medicine Bow Mountains of Wyoming. The later peak in moth observations here at higher elevations is consistent with the earlier peak (in red) at the low-elevation site farther east.
Figure B shows data on moth abundance collected by Don White at lower elevations just east of the Rocky Mountains (light turquoise dots: Plains) and at moth aggregation sites in alpine areas to the west (dark blue dots: Peaks). All of these data suggest that moth aggregations culminate by mid-July and last until early September, which is consistent with more casual observations of moth abundance by Sean O'Brien in the Absarokas (the red band denoting "Abundant moths" in A), as well as peak levels of bear feeding shown as the vertical blue shaded band and data in Figure D.
Bear activity on moth sites the Absaroka Mountains unambiguously reflects moth phenology. Bear activity is at its peak from around mid-July to early September, centered on around August 18th, coincident not only with peak moth abundance, but also the culmination of fat accumulation by female moths at between 70 and 80%. Trends in fat content of both male and female moths are shown in figure C, based on data from multiple sources; data on bear activity come from ground observations by Steve and Marilyn French, as well as aerial observations by the Interagency Grizzly Bear Study Team.
Secondarily, sex, age, and reproductive composition of bears on moth sites tends to vary during the summer and early fall, at least in the Absaroka Mountains. Figure E shows the composition of bears observed on moth sites around Yellowstone (i.e., the Absarokas) as well as in Glacier National Park, with the proportion comprised of family groups shown in green. Composition of bears on moth sites in Glacier appeared to be relative stable, whereas the proportion of bears comprised of females with cubs or yearlings in Yellowstone appeared to decline from a peak in late June to a low during late August and early September...for reasons that are not readily explicable.
Even so, family groups comprise a larger proportion of total observed bears than would be expected by their numbers in the population, by about 20%, and lone adults a smaller fraction, by about 17%. These patterns hold for multiple studies and study areas, including the Mission Mountains of western Montana, Glacier National Park, and the Absarokas, although all of the data I use were collected several decades ago. It is possible--perhaps likely--that the pattern has changed during recent years. But it is probably true that moths were a disproportionately important source of energy for reproductive females in areas where grizzlies exploited moths and, moreover, an unambiguously high-quality food (see this entry on bear nutrition). There is no other item in grizzly bear diets anywhere with a fat content as high as that of moths during late summer and early fall--coincident with the period when bears themselves are accumulating most of the adipose reserves that will see them through the winter and following spring (see this entry on life strategy of grizzlies).
Trends in Daily Activity
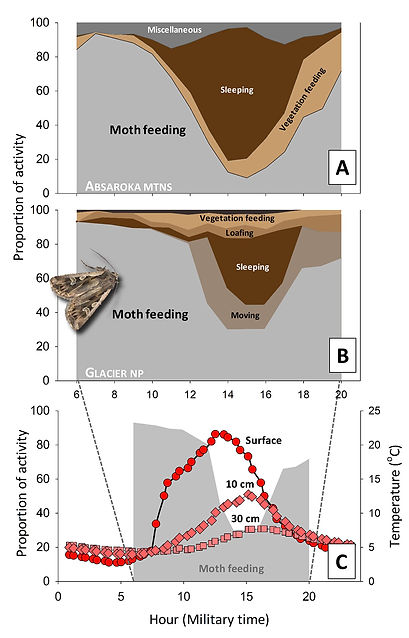
Trends in the daily activity of bears on moth sites mirror trends in activity of moths, which mirror, in turn, predictable trends in not only the ambient environment, but also the environment of talus refugia. Deborah Kendall and Peter Kevan perhaps most thoroughly documented daily activity patterns of cutworm moths in alpine habitats, albeit farther south than where documented bear feeding occurs, in central Colorado. They conclusively documented peak foraging on nectar by moths at night, but beginning in early evening after daytime temperatures cooled, and ending during early morning as temperatures rose. Both Don White and Sean O'Brien documented much the same pattern farther north.
By all indications, though, moths don't stop managing their environment once they enter the cooler nether-regions of alpine talus. As the surface layers gradually heat, moths make their way progressively deeper and, in the process, maintain their environment at around 5-10 degrees C (40-50 degrees F). Cutworm moths are remarkably cold tolerant and surprisingly vulnerable, physically, to high temperatures. It is not clear why moths go to such extremes to sequester themselves in the comparative cold, but I speculate it has something to do with escaping parasitic wasps and flies, which are the moth's primary predators at lower elevations.
Feeding on moths by bears peaks during morning and evening hours when moths are leaving or entering talus refugia, and at the shallowest depths within. This crepuscular pattern of peak feeding on moths is clearly evident in activity patterns of bears documented by Sean O'Brien in the Absarokas (A at right) and Don White in Glacier (B). The other dominant activity is sleeping, which is concentrated during the middle of the day while moths are deepest in the talus. Otherwise, bears are not doing much else. Figure C, comprised of temperature data that Don collected in Glacier, simply registers bear feeding against temperature profiles at different depths, showing progressive surface heating and stable temperatures at depth, with peak differential at around 3 pm (15 horus military time), at the same time that moths are presumably deepest and level of bear feeding least. Which all makes sense.